utf-8МОСКОВСКАЯ СЕЛЬСКОХОЗЯЙСТВЕННАЯ АКАДЕМИЯ
РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ
ЗООИНЖЕНЕРНЫЙ ФАКУЛЬТЕТ
______________________________________________________________________
ДИПЛОМНАЯ РАБОТА - УСТАРЕЛА. СМОТРИТЕ НОВУЮ РЕДАКЦИЮ http://orlik62.narod.ru/links_diplom.html
На тему: “Анализ соответствия традиционных технологий быстроаллюрного коневодства России современным зоотехническим представлениям”
Выполнил студент 51 группы вечернего обучения зоотехнического
факультета Дёмин И.И.
Руководитель – к.с/х.н. Парфенов В.А.
Москва – 2005
СОДЕРЖАНИЕ
Введение
Глава 1. Влияние экологических факторов на воспроизводительную функцию племенных быстроаллюрных лошадей.
Глава 2. Технологии выращивания и содержания быстроаллюрных лошадей.
2.1. Влияние условий среды и физической нагрузки на организм животных
2.2. Технология выращивания жеребят быстроаллюрных пород
2.3. Эксплуатация, тренинг быстроаллюрной лошади и организация моциона племенных быстроаллюрных лошадей
Глава 3. Организация кормления лошадей
3.1. Организация обеспечения всех половозрастных групп лошадей полноценными сбалансированными физиологически обоснованными кормами
3.2. Значение минеральных веществ в кормлении лошадей
3.3. Значение витаминов в кормлении лошадей.
3.4. Протеиновая питательность кормов и баланс аминокислот. Значение баланса аминокислот для различных половозрастных групп быстроаллюрных лошадей.
3.5. Углеводная питательность корма и значение биологических экстрактивных веществ
3.6. Липидная питательность кормов и значение жиров в жизни быстроаллюрных лошадей
3.7. Энергетическая ценность кормов
3.8. Характеристика основных кормов
3.8.1. Сено и солома
3.8.2 Перспектива применения сенажа
3.8.3. Корне- и клубнеплоды. Фрукты.
3.8.4. Веточный корм.
3.8.5. Злаковые и бобовые корма.
3.8.6. Комбикорма неполнорационные, полнорационные и условно полнорационные.
3.9. Организация прочной кормовой базы
3.10. Анализ сбалансированности по питательным веществам традиционных рационов быстроаллюрных лошадей
.
3.11. Разработка подкормки для жеребят-сосунов рысистых пород лошадей
Глава 4. Механизация производственных процессов на коневодческих предприятиях
4.1. Обеспечение комфортных условий содержания и тренинга быстроаллюрных лошадей
Глава 5. Профилактические ветеринарные мероприятия и зоогигиена
5.1. Обеспечение благополучной ветеринарно-санитарной обстановки
5.2. Значение борьбы с заразными инфекционными болезнями.
5.2.1. Вирусный артериит.
5.3. Значение противопаразитарных мероприятий
5.4. Зоогигиена и профилактика
Глава 6. Организационно-правовые мероприятия организации коневодческого дела
6.1. Рациональная расстановка, воспитание и подготовка кадров коневодства
6.2. Создание материальной и моральной заинтересованности в продолжительной и добросовестной работе.
6.3. Племенное дело
6.3.1. Значение достоверности информации используемой в племенной работе.
6.3.2. Оценка интерьера. Система модельных характеристик интерьера быстроаллюрной лошади.
6.3.3. Эффект гетерозиса и спортивное коневодство.
6.4. Организация спортивного коневодства.
6.5. Зоотехнический аудит и лицензирование в конном деле.
6.5.1. О необходимости введения в практику животноводческой отрасли зоотехнического аудита.
6.5.2. Зоотехнический аудит – организационно-экономический инструмент управления животноводческой отраслью.
6.5.3. Общая модель зоотехнического аудита
6.5.4. Общие принципы аудита систем экологического управления и менеджмента.
6.5.5. Алгоритм управления зоотехнической аудиторской деятельностью.
6.5.6. Направление применения результатов зоотехнического аудита
6.5.7. Применение зоотехнического аудита в быстроаллюрном коневодстве.
Заключение
Приложения
Литература
Введение
Основные приемы зоотехнии начали зарождаться еще при первобытнообщинном строе, одновременно с возникновением животноводства. За всю историю доместикации человек предъявлял животным различные требования, которые произрастали из его собственных потребностей. Животноводство служит человеку источником высокоценных продуктов питания (молоко, мясо, яйцо и т.п.), сырья для промышленности (шерсть, мех, кожевенное сырье, пух и т.п.) и энергии для вспашки, перевозки грузов и т.п.
На первом этапе человек был озадачен выбором животных с полезными для него отклонениями признаков, закреплением этих выбранных отклонений в потомстве путем отбора в производящий состав лучших животных.
В последствие в зоотехническую культуру влилось понятие подбора, которое вместе с отбором и подразумевают целенаправленную деятельность человека по совершенствованию наследственного потенциала полезных признаков сельскохозяйственных животных. Отбор и подбор и по ныне являются сутью селекции.
Накопив определенный потенциал знаний, человек сформулировал для себя ещё более широкое понятие – племенная работа. Значение нововведения состояло в том, что наследственные задатки хозяйственно-полезных признаков животные способны проявлять только в определенных условиях внешней среды, названных человеком – комфортными. С тех пор в понятие “племенная работа” вкладывается смысл не только “собственно селекция”, но и обеспечение этой работы комфортными условиями внешней среды (1), а также физиологически обоснованной организацией кормления, при ведении селекции в популяции здоровых животных с обязательным соблюдением санитарно-ветеринарных требований к условиям содержания.
Однако даже при полном понимании значимости каждой составной части, племенное дело было поставлено человеком себе на службу не одинаково в различных отраслях животноводства. К настоящему моменту наибольшего расцвета племенная работа достигла в птицеводстве, аквакультуре и свиноводстве, а наименьшего, по нашему мнению, в овцеводстве и коневодстве.
На протяжении уже 6 тысяч лет лошадь является верным спутником человека. На протяжении этого времени роль лошади в социально-экономической сфере человеческой цивилизации изменялась в соответствии с изменениями уклада жизни человека (2). Значение лошади в жизни человека обеспечивалось универсальностью её использования, высокая интенсивность роста молодняка, низкая себестоимость работ и продукции, высокая биологическая ценность конины и молока.
В эпоху научно-технического прогресса машины заменили лошадей во многих областях, тем не менее, интерес к этим животным не ослабевает до сих пор.
В течение нескольких десятилетий мировое поголовье лошадей зафиксировалось на уровне 65 миллионов голов (3). По мере того, как меняется жизнь человека, изменяется и интерес человека к лошади. Наиболее динамично лошадь “завоевывает сферы спорта, развлечений, активного отдыха, восстановления здоровья людей, производства уникальных продуктов питания” (3). Ипподромный бизнес, как самостоятельная отрасль, но опирающаяся на скаковое и рысистое коневодства, достиг денежного оборота в десятки миллиардов долларов ежегодно.
Однако, несмотря на структурную перестройку коневодства и высокую динамику освоения новых сфер деятельности лошадьми, технологии разработанные человеком в период от начала доместикации до момента создания чистокровной верховой породы, к настоящему времени мало изменились.
В птицеводстве, аквакультуре и свиноводстве в полном объеме пользуются преимуществами комфортных условий содержания животных, широко применяется промышленное скрещивание. В тоже время в коневодстве этим приемам зоотехнии уделяется недостаточное внимание. Усугубляется это обстоятельство ещё и тем, что северные и центральные регионы России с их климатическими условиями, относятся к зонам рискового животноводства, по причине значительного влияния внешней среды на хозяйственно-полезные качества выращиваемых животных на пастбищах и в табунах.
Обзорно-аналитический характер данной дипломной работы позволяет сопоставить соответствие традиционных технологий в быстроаллюрном коневодстве России научным знаниям в области зоотехнии на современном этапе.
В данной дипломной работе в качестве практического приложения приводится разработка сбалансированной подкормки жеребят-сосунов рысистых пород лошадей. Также приводится разработка концептуальных основ зоотехнического аудита.
Глава 1. Влияние экологических факторов на воспроизводительную функцию племенных быстроаллюрных лошадей.
Основное условие, определяющее динамику развития коневодческой отрасли – организация расширенного воспроизводства конского поголовья (2).
Длительное время доминировал подход к изучению влияния факторов внешней среды на лошадей в совокупности со стремлением максимального снижения себестоимости выращиваемых лошадей. По-нашему мнению, данный подход может широко применяться для продуктивного и рабочепользовательского коневодств, основанных на экстенсивных табунно-пастбищных технологиях и совершенно недопустимых для быстроаллюрного коневодства.
Воспроизводство лошадей складывается в основном из пяти основных периодов – оплодотворения кобыл, их жеребости, выжеребки, выращивания молодняка и выдержки племенной быстроаллюрной лошади. Все эти стадии требуют глубоких знаний физиологии содержания, тренинга, кормления и ухода за лошадьми, умения грамотного применения этих знаний для формирования эффективных технологических процессов на основе современных представлений о приемах и способах животноводства.
В комфортных условиях содержания, полноценного и сбалансированного кормления, реализуются заложенные в зиготе наследственные свойства родительских пар. В таких условиях влияние внешней среды на организм животного минимальный и процессы роста и развития формируют организм животного максимально соответствующий его геному.
Содержанию и кормлению кобыл в период беременности человек старается уделять большое внимание. Однако очень редко человек уделяет внимание экологии и не только содержанию в корме, кормовых добавках и воде загрязняющих веществ (нитраты, удобрения не биологического происхождения, ядохимикаты, гербициды, тяжелые металлы, нефтепродукты, консерванты, красители и т.д.), но и содержанию вредных веществ в окружающей жеребую лошадь среде.
При действии на организм животного неблагоприятных факторов, проявляется морфо-физиологическая адаптация, проявляющаяся в координации приспособительных процессов при помощи двух главных систем – нервной и гуморальной.
Общеизвестно, что любое живое существо, приспосабливается к условиям своего обитания: сезонные явления, температурный режим, влажность, давление, солнечная радиация, состав пищи, воды, воздуха и др. При воздействии на организм животного извне любых химических веществ возникает ответ – синтез соответствующих гормонов и ферментов, необходимых для обеспечения усвоения, путем трансформации в доступные для потребления организмом формы. Приспособление к химизму и характеру внешних условий отражается в устройстве систем потребления и обмена с внешней средой (пищеварение,
дыхания, терморегуляции, выделения и т.п.), а также выражается в иммунных реакциях организма и составе иммунного фона. При попадании в организм нового вещества, не появлявшегося ранее, начинается синтез новых форм ферментов, а также изменяются гормональный фон и иммунные реакции. Таким образом, любые качественные и количественные изменения состава веществ, поступающих из внешней среды, неминуемо приводят к генетически обусловленного состава ферментов, гормонального и иммунного фонов организма.
Из эмбриологии и иммунологии известно, что между организмом матери и плодом существует иммунологическая несовместимость. Плод, имеющий геном, отличный от материнского генома (на 50%), является для неё чужеродным. По этой причине, между ними всегда существует иммунный конфликт, подавляемый механизмом, способствующим сохранению плода и препятствующим его отторжению. Плацента и плод синтезируют ряд белковых факторов и гормонов (эстрогены, простагландины, прогестерон), подавляющие реакцию отторжения (6).
При попадании в организм жеребой кобылы новых веществ или их нового количества, её организм адаптируется к новым веществам, а вот плод этой возможности практически не имеет. В результате биохимической адаптации кобылы может возникнуть тканевая несовместимость с неадаптированным плодом. В результате иммунологической атаки на плод нарушаются условия его питания, роста и развития, вследствие чего могут возникнуть отклонения в его развитии, уродство и аллергия(6).
Для убедительности и конкретизации вышеизложенного приведем ниже информацию о некоторых проведенных исследованиях.
Так было получено значительное количество подтверждающих данных о том, что замедление развития плода в матке является очень важным фактором риска возникновения сердечно-сосудистых нарушений и нарушения обмена веществ во взрослом возрасте. Эксперименты же, проводимые на животных (морские свинки, крысы, овцы), подтвердили взаимосвязь между задержкой внутриутробного развития и формированием артериальной гипертензии или обменных нарушений во взрослом возрасте. Независимо от причины, приводящей к нарушению внутриутробного развития у животных (нарушение маточного кровотока, анемия у матери, недостаточное калорийное питание, либо низкобелковая диета во время беременности), результат получался один и тот же (6).
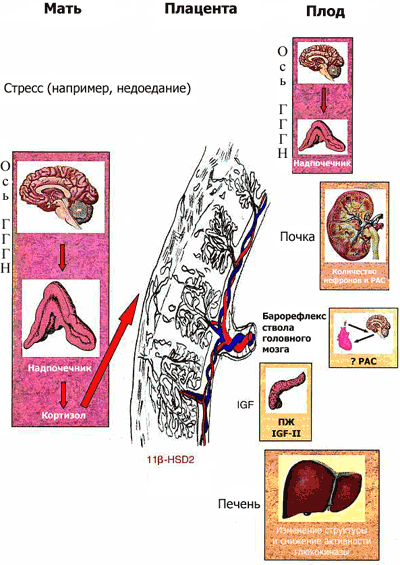
Рис. 1 Потенциальные механизмы "программирования плода"
Стресс материнского организма (например, недоедание во время беременности) приводит к активации оси гиппокамп-гипоталамус-гипофиз-надпочечники (ГГГН) у беременной, что, в свою очередь, в зависимости от уровня активности фермента плаценты, инактивирующего кортикостероиды (11 -гидроксистероиддегидрогеназы 2-го типа, 11 HSD-2), приводит к воздействию на плод высоких концентраций глюкокортикоидов (ГК). Высокий уровень глюкокортикоидов в крови плода способствует изменению числа стероидных рецепторов в ГГГН системе, изменяет уровень активности других функциональных систем (гормон роста /инсулин-зависимый фактор роста,
ренин-ангиотензиновая система, вызывает замещение - клеток поджелудочной железы плода - клетками, характерными для взрослого организма), что в конечном итоге приводит к патологическим изменениям во взрослом возрасте. IGF II: инсулиноподобный фактор роста II; РАС: ренин-ангиотензиновая система; ПЖ: поджелудочная железа (6).
Потомство этих животных имело:
- существенные изменения в морфометрии печени и значительное снижение активности глюкокиназы в перивенозных областях (у крыс);
- артериальную гипертензию, уменьшение объема мышечной ткани и увеличение жировой ткани, нарушение толерантности к глюкозе, нарушенный гомеостаз холестерина (у морских свинок);
- артериальную гипертонию к 3-х месячному возрасту (у овец) (6).
Имеются также неопровержимые данные о том, что реализация генетической программы формирования головного мозга осуществляется во взаимодействии со средовыми факторами. Эмбриональный и ранний постэмбриональный периоды развития животного играют исключительно важную роль в формировании головного мозга
.
Рост и развитие мозга может нарушаться под влиянием ряда факторов:
- высокой чувствительностью мозга в критические периоды развития к внешним воздействиям,
- последствиями воздействий (нейро-гуморальные, токсические, физические, медикаментозные и т.п.), имеющими необратимый характер, как постоянно действующие факторы.
Необратимость последствий воздействий зависит, прежде всего, от того, что зрелые нервные клетки (нейроны) не обладают способностью к делению, в связи, с чем любые факторы, которые приводят к уменьшению их численности к моменту рождения, делают мозг малонейронным, вследствие, уменьшения интенсивности деления нейробластов или увеличение числа погибающих нейробластов и нейронов, с автоматическим отклонением от нормального процессов формирования связей между нейронами - образования синапсов (7).
В исследуемых литературных данных указано на то, что у разных линий крыс животные с большей массой мозга обладают лучшими способностями к обучению. Мыши с большей массой головного мозга быстрее достигают критерия обученности и успешнее обучаются в дальнейшем (7).
Также обнаружена и закономерность, что факторы, приводящие к уменьшенной массе тела, вызывают уменьшение массы головного мозга, а также установлена положительная корреляционная связь массы тела и мозга у новорожденных, которая проявляется как у человека, так и у экспериментальных животных (7).
Причинами низкой массы тела при рождении указываются качественная и количественная неполноценность питания матери во время беременности, многоплодная беременность, различные болезни матери, нарушающие развитие плаценты, снижающие ее транспортные функции. Во всех описанных ситуациях плод испытывает дефицит питательных веществ, который может снижать интенсивность синтетических процессов в нейронах, скорость роста их отростков. Кроме того, к снижению массы тела плода могут вести также воздействия радиации, алкоголя, наркотиков, лекарственных препаратов, вирусов и бактерий. Эти агенты могут приводить к поражению плаценты, уменьшать ее способность доставки необходимых компонентов от матери к плоду и удаления шлаковых продуктов последнего, то есть эффект их воздействий также может реализовываться через уменьшение количества поступающих питательных веществ. Перечисленные факторы обладают способностью и прямо повреждать клетки развивающегося мозга. Исследованиями в области гормонального фона установлено, что гормоны, циркулирующие в крови плода, синтезируются плацентой, эндокринными железами матери и его собственными железами. Рост, развитие и функционирование желез внутренней секреции уже в эмбриогенезе в значительной степени регулируются гипоталамусом (7).
Также установлено, что гормональные воздействия оказывают:
- необратимое, программирующее, изменяющее фенотип и функциональную активность клеток влияние,
- обратимое, регуляторное, изменяющее клеточные функции (7).
Влияние концентрации гормонов в крови матери на мозг плода реализуется различными путями:
- часть гормонов (стероидные), продуцируемые корой надпочечников, яичниками, проходит через плаценту в кровь плода, где они прямо воздействуют на нейробласты, нейроны и другие клетки мозга;
- гормоны гипофиза, яичников, надпочечников и других желез матери влияют на развитие плаценты, определяя эффективность транспорта различных компонентов от матери к плоду и в обратном направлении;
- изменения эндокринных функций плаценты, вырабатывающей большое число различных гормонов, поступающих как к матери, так и к плоду, также могут оказать воздействие на формирование мозга;
- изменения содержания гормонов в крови матери и плода влияет на развитие его мозга и опосредованно, воздействуя на концентрацию в крови таких важных для этого процесса веществ, как глюкоза, аминокислоты, разные классы липидов и т.д.;
- при снижении функции желез у матери железы плода, стремясь компенсировать дефицит соответствующих гормонов, находятся в состоянии гиперфункции, а следствием этих процессов является то, что после рождения характер развития соответствующих желез, их функциональная активность, реакция на стимулирующие воздействия отличаются от контрольных, что немаловажно, в части показателей высшей нервной деятельности у животных с измененными вследствие указанных выше причин эндокринными железами (7).
Приняв к сведению вышеизложенное, рассмотрим некоторые фрагменты существующих ныне технологий, сопряженных с процессами воспроизводства быстроаллюрных лошадей.
“Случку при конюшенном содержании проводят с 15 февраля по 15 июля, при табунном – с 15 апреля по 1 августа. Эти сроки могут несколько изменяться в зависимости от климатических условий. Во всех хозяйствах целесообразно проводить случку кобыл, по возможности, в более ранние сроки, чтобы окончить выжеребку до начала весенних полевых работ и чтобы жеребята к отъему нормально развились” (5-стр.99).
По нашему мнению, зависимость технологии современного быстроаллюрного коневодства от климатических и иных сезонных факторов представляет серьезное препятствие по организации интенсивного и расширенного воспроизводства конского поголовья из-за частых прохолостов кобыл, возникающих по различным причинам и как следствие этого – понижение зажеребляемости и выхода жеребят. Сезонность выжеребки в некоторой степени также оказывает влияние на сезонную потребность, определенными технологиями конезаводства, в квалифицированных кадрах. По этим причинам, снятие воздействия сезонных факторов на современные технологии коневодства - одна из проблем, требующих своего инженерно-технического и организационно-правового решения.
Инженерно-технические решения, по нашему мнению, должны акцентироваться на вопросах экологии содержания, тренинга и кормления лошадей. В то время как организационно-правовые решения должны направляться на снятие каких-либо регламентирующих, как и других фактически существующих ограничений, обусловленных сроками выжеребки быстроаллюрных лошадей.
Одним из заблуждений, доставшихся нам из прошлого, представляется нам недооценка значимости маточного поголовья в сравнении с жеребцами-производителями, что неминуемо отражается на качестве формируемого маточного состава. В доказательство данного опасения приводим некоторые выдержки из руководств.
“Матки из-за малоплодности не могут оказать такого влияния на структуру пород, как производители...” (2).
“Согласно наблюдениям, жеребчики более требовательны к условиям кормления и содержания, чем кобылки, и значительно лучше отзываются на полноценное кормление и хорошее содержание.… Поэтому нормы кормления жеребчиков должны быть несколько выше норм кормления кобылок” (4-стр.122).
По нашему мнению, различия в нормах кормления жеребчиков и кобылок не означают большую или меньшую требовательность к качеству рационов, а свидетельствуют о количественной разнице в потребностях, обусловленной физиологическими различиями в росте и развитии, и никаким образом не означает различия в прихотливости. А вместе с тем степень развития будущих племенных кобыл, куда сильнее и глубже может повлиять на качество их будущего приплода, множеством факторов (см. выше), нежели степень развития будущих жеребцов-производителей.
Жеребость – сложный физиологический процесс, требующий высококвалифицированного подхода для обеспечения правильных условий кормления, содержания и проведения моциона кобыл.
Таким образом, влияние условий содержания племенных лошадей имеет качественную и количественную составляющие.
Качественная составляющая заключается в улучшении состояния здоровья получаемого приплода, а также улучшении показателей развития и роста жеребят и как следствие, возможности выращивания более ценного поголовья быстроаллюрных лошадей.
Проблематика коневодства, связанная с периодом развития и роста жеребят, требует от зоотехнических специалистов комплексного подхода, выражаемого в научно обоснованных и ясных решениях. Под ясностью решений нами подразумевается, прежде всего, однозначность и понятность способов достижения цели, поставленной перед технологией её преследующей.
Далее приведем выдержку о том, как предлагается понимать решение задачи кормления жеребых кобыл применительно к существующей практике.
“Забота о жеребенке должна начинаться, когда он еще в утробе матери. Жеребых кобыл обеспечивают полноценными кормами: сеном, концентратами, корнеплодами, кормами, богатыми витаминами (особенно каротином) и минеральными веществами. В первой половине жеребости, когда формируются ткани и органы зародыша, неполноценное питание кобыл, прежде всего недостаток в кормах витаминов и микроэлементов, часто приводит к скрытым абортам. Во второй половине жеребости в развивающемся плоде интенсивно нарастает масса скелета; у новорожденного жеребенка она составляет 31% его живой массы. Резко увеличивается потребность жеребой кобылы в кальции и фосфоре. В последние месяцы жеребости усиливается нарастание мышц плода и увеличивается потребность в белковых веществах, поэтому так важно полноценное протеиновое кормление” (5- стр.104).
По нашему мнению, термин “полноценный корм” означает покрытие потребностей организма необходимым количеством переваримых веществ, содержащихся в корме. Однако, применение полноценного корма не равнозначно с понятием комфортного кормления, прежде всего ввиду того, что комфортное кормление предполагает ещё, сбалансированное кормление и даже не только это. При этом надо ещё учитывать степень эксплуатации (интенсивность работы) лошади. В свою очередь, степень эксплуатации определяется величиной селекционируемого человеком хозяйстсвенно-полезного признака в заданный момент времени и в конкретных условиях. Таким образом, рацион племенной быстроаллюрной лошади должен быть обеспечен, как переваримыми веществами в свободном и связанном виде, так и непериваримыми (балластными), в количествах минимально возможных, достаточных и необходимых для обеспечения всех трофических потребностей организма лошади от одного приема корма до другого с учетом индивидуальных особенностей животного, в том числе времени необходимого до начала поступления в кровь лошади новой порции питательных веществ от новой порции корма и т.д.
По нашему мнению, принцип сбалансированности и адекватности, соответствующему режиму продуктивности животного, корма для удобства реализации в конкретных рационах, необходимо дифференцировать по группам питательных веществ, разделенных по степени значимости для обеспечения трофических потребностей:
- основные питательные вещества (переваримые протеин, жиры, углеводы);
- протеин (соотношение аминокислот
);
- углеводы (соотношение БЭВ, легко- и труднопереваримой клетчатки);
- жиры и масла (соотношение жирных кислот);
- минеральные вещества (соотношение макро- и микроэлементов);
- биологически активные вещества (соотношение витаминов).
Уровень организации кормления во многом определяется и квалификацией обслуживающего лошадей персонала.
Учитывая узкий набор кормовых средств, традиционно применяемых для кормления лошадей быстроаллюрных пород (овес, сено, шрот, меласса, сахар, травяная мука) и невозможность замены сбалансированного рациона кормовыми добавками, которые не решают проблемы лишних (а часто и вредных из-за переизбытка) веществ и понижают общую переваримость рациона, можно утверждать, что кормлению лошадей повсеместно уделяется неоправданно мало внимания.
Глава 2. Технологии выращивания и содержания быстроаллюрных лошадей.
2.1. Влияние условий среды и физической нагрузки на организм животных
Способность живых существ приспосабливаться к меняющимся условиям среды – одна из их важнейших особенностей, позволяющих организмам существовать в самых различных условиях внешней среды.
Здоровье лошади зависит, главным образом от условий во внешней и внутренней средах, факторы и свойства которых, воздействуя на организм лошади в качестве раздражителей, определяют естественную реакцию организма лошади на эти воздействия, и последующую адаптацию к данным условиям.
Лошади, как и многие другие животные, дышащие атмосферным воздухом, находятся в постоянном взаимодействии с воздушной средой. По мере эволюции вида, природа выработала у лошадей ряд качеств, отвечающих на воздействие факторов окружающей среды. Так температуре окружающей среды противостоит терморегуляция, а влажности воздуха – транспирация и перспирация.
К наиболее важным факторам окружающей среды принято относить следующие:
1. Температура воздуха;
2. Газовый состав воздуха;
3. Влажность и скорость воздуха;
4. Давление воздуха;
5. Аэроионизация;
6. Пылевая и микробная загрязненность воздуха;
7. Солнечная радиация;
8. Электрические и электромагнитные поля;
9. Шум.
В процессе эволюции видов в каждой климатической зоне появляются виды животных соответствующих именно данному набору параметров среды их обитания.
Так в жарком климате у лошадей тонкая кожа с редкими, короткими и тонкими волосами, слаборазвитая подкожная клетчатка, сухой тип конституции, слаборазвитые челка, хвост и щетки. В тоже время лошади северной зоны имеют ярко выраженную способность к быстрому жироотложению (2).
Температурная адаптация лошадей сопровождается совокупностью специфических физиологических изменений, главными из которых, по мере пребывания в условиях повышенной температуры среды, являются усиление потоотделения, снижение температуры ядра и поверхности тела, уменьшение частоты сердечных сокращений при нагрузке и увеличение числа секретируемых потовых желез. Механизмы температурной адаптации направлены на усиление отдачи тепла организмом лошади во внешнюю среду. При любом увеличении температуры организма лошади, в частности и во время работы, потоотделение начинается быстрее, т.е. снижается температурный порог потоотделения и увеличивается его скорость (35).
После адаптации организма к жаре меньше пота стекает в виде капелек, не испаряясь, так как пот более равномерно распределяется по всей поверхности тела, чем у неадаптированного животного, т.е. возрастает площадь поверхности кожи, охваченной для усиленной теплоотдачи испарением. Как следствие, усиление потоиспарения ведет к более эффективному снижению температуры кожи, благодаря чему кровь кожных покровов охлаждается быстрее, усиливается физический транспорт тепла от глубоких частей тела и более нагретых к его поверхности (35).
Главным эффектом усиления адаптивных температурных механизмов является снижение температуры тела, как в условиях покоя, так и её прирост в момент мышечной работы (35), увеличивается температурный диапазон мышечной работы, а также снижается тепловая нагрузка в легких из-за снижения суммарного тепла реакций оксигенации, что в свою очередь ведет к увеличению коэффициента полезного действия (к.п.д.) мышечных волокон.
В течение температурной адаптации снижается содержание солей в поте, увеличивается выделение воды и как следствие, концентрация электролитов в крови повышается, что рефлекторно вызывает сильное ощущение жажды, являющееся механизмом, направленным на компенсацию потерь воды организмом животного. У неадаптированного к жаре организма данный рефлекс хуже выражен, что может приводить к обезвоживанию организма, а при резком водопотреблении и к опою. Адаптированное к жаре животное имеет четко выраженный рефлекс для поддержания водного баланса организма (35).
По мере адаптации к высокой температуре снижается проницаемость капилляров кожного покрова, уменьшается выход молекул белка из капилляров, содержание белка в тканевой жидкости кожи увеличивается. При тепловых воздействиях белок интенсивно перемещается транзитно через лимфатическую сеть кожи в кровь, что позволяет сохранять её высокие онкотическое давление и объем. Объем циркулирующей крови в состоянии покоя увеличивается, а показатель гематокрита и вязкость крови имеют тенденцию к снижению (35).
Температурная адаптация приводит к снижению нагрузки на сердечно-сосудистую систему, к росту возможности эффективного усиления кожного кровотока, за счет более быстрого перемещения крови в систему кожных капилляров, а за счет раскрытия и расширения сети поверхностных капилляров, к приближению крови к поверхности тела животного и более эффективному её распределению (35).
По мере протекания процесса температурной адаптации уменьшается степень сужения сосудов в чревной и почечной областях, что улучшает кровоснабжение органов брюшной полости во время физической работы, повышается механическая эффективность выполнения физической работы в условиях повышенной температуры, сопровождаемая прогрессивным снижением потребления кислорода при выполнении работы, снижается частота сердечных
сокращений в покое и при работе, увеличивается систолический объем, что не изменяет сердечный выброс на протяжении периода пребывания в жарких условиях, а также увеличивается венозный возврат, за счет повышения объема циркулирующей крови (35).
Тренировочные и состязательные нагрузки, требующие проявления выносливости, вызывают существенное повышение температуры ядра тела – до 40
оС, даже в нейтральных условиях среды, что служит стимулом для развития адаптационных реакций к повышенной тепловой нагрузке с морфологическим оформлением ответа на эти реакции. Данные реакции, со стороны сердечно-сосудистой системы, потовых желез и других органов, во многом сходны с реакцией организма животного на акклиматизацию к жарким условиям (35).
В результате систематических физических нагрузок у тренируемых животных совершенствуется терморегуляция:
- снижается теплопродукция;
- повышается чувствительность реакции потоотделения на тепловое раздражение, выражающееся в сокращении времени от начала теплового воздействия до начала потоотделения;
- увеличивается объем циркулирующей крови;
- совершенствуются реакции перераспределения кровотока за счет раскрытия-сужения сосудов и подключения в работу закрытых сосудов кожного покрова;
- более равномерное распределение потоотделения по поверхности тела, за счет увеличения числа функционирующих потовых желез;
- понижается концентрация солей в поте;
- улучшается способность к теплопотерям за счет повышенного потообразования;
- понижается внутренняя и кожная температура тела (35).
Вышеизложенное морфологическое оформление ответа на реакции к повышению температуры и к физической нагрузке особенно важно учитывать с момента рождения жеребенка и последующей адаптации к условиям внешней среды, при этом важным моментом совершенствования его организма в связи с последующей специализацией использования, является как возможно ранняя организация тренировочного процесса с целью грамотного задания адекватной степени развития организма жеребенка физической нагрузки.
Оптимальному значению каждой из переменных внутренней среды организма (температура тела, рО
2 в тканях и т.п.) соответствуют диапазоны значений внешних факторов природы. В случае отклонения от этих значений включается физиологическая регуляция, направленное на поддержание гомеостаза, на расширение и смещение диапазона устойчивости организма. Главенствующее положение в системе адаптации организма занимает установление оптимизированного стабилизированного уровня затрат энергии и в связи с этим, снижение энергетической цены гомеостаза становится основным принципом физиологической адаптации к воздействию внешних факторов.
В качестве естественных разобщителей окисления и фосфорилирования выступают тиреоидные гормоны, повышающие теплопродукцию мышечного сокращения (24-стр.128).
При хроническом действии низкой температуры в мышцах происходит увеличение относительного количества медленных (красных) и промежуточных волокон, для которых характерен изометрический тип мышечных сокращений, а также происходит повышение потребления медленными (красными) волокнами кислорода, что усиливает гипоксию при двигательной деятельности (15-стр.116). Кроме того, при длительном воздействии низких температур происходит угнетение функций физической терморегуляции, что одновременно понижает эффективность мышечной работы (к.п.д. мышц), которая напрямую зависит от эффективности системы теплоотдачи (системы “охлаждения” организма).
К.П.Д. мышц – представляет собой отношение кинетической энергии мышечного сокращения ко всему количеству энергии, вырабатываемой мышцами по время своей работы. Постепенно, к.п.д. тренированной мышцы повышается и может достигать 50% (18-стр.17).
В условиях покоя мышцы потребляют 20-25% всей энергии, которую производит организм. В обычных условиях мышечная масса вырабатывает 50-60% тепла организма. При усиленной работе этот показатель достигает 80%, поэтому эволюцией видов мышцам отведена ещё одна важнейшая функция – участие в терморегуляции (18-стр.11). Во время работы одна часть энергии превращается в тепло, а другая – преобразуется в механическую работу, при этом соотношения между этими частями отражает к.п.д. мышечных сокращений. При этом, учитывая тот факт, что сократительный белок не может выполнять свою функцию в условиях денатурации, к.п.д. мышц отчасти отражает уровень готовности мышечной ткани к выполнению физическую нагрузки максимальной мощности.
В процессе эволюции гомойотермные животные выработали способность использовать двигательную активность для защиты от холода, как во взрослом состоянии, так и в новорожденном, когда эта способность наиболее ярко выражена. На основании литературных данных и по научным исследованиям сделано утверждение, что “… все случаи химической терморегуляции сочетаются с повышением сократительной активности мышц. Это – двигательная активность, холодовая дрожь, терморегуляционный тонус. Мощность их довольно велика” (17-стр.39).
Также было установлено, что при акклиматизации к холоду химическая терморегуляция осуществляется при низком уровне электрической активности мышц. Это навело ученых на мысль, что “… после акклиматизации к холоду теплопродукция мышечных сокращений существенно повышается” (17-стр.40) и этот факт был подтвержден прямым определением потребления кислорода мышцами бедра акклиматизированных и контрольных кроликов. Исследования, проведенные на изолированной диафрагме крыс, показали увеличение теплопродукции, рассчитанной на 1 г массы и 1 г развиваемой силы, у акклиматизированных к холоду животных на 38% и 54% соответственно больше, чем у контрольных (17-стр.41). Что касается механизма уменьшения к.п.д. мышц, то
учеными-исследователями делается вывод,
“…что множество даже очень слабых сокращений, которые не производят практически никакой механической работы, могут стать источником значительного количества тепла мышечной ткани. Но именно такая деятельность имеет место в холодовой мышечной дрожи или при так называемом терморегуляционном мышечном тонусе, состоящих из частых, слабых, нерегулируемых сокращений отдельных групп мышечных волокон” (17-стр.45),
а механизмом понижения к.п.д. мышечной деятельности и повышения теплопродукции может являться разобщение окисления и фосфорилирования (17-стр.46).
Относительно уровня понижения к.п.д. мышц, исследованиями на мышцах задних конечностей кролика установлено, что после акклиматизации к холоду к.п.д. мышц падает в
1,5 – 2 раза, при этом эффективность терморегуляционного тонуса повышается на 200-300%, а прирост теплопродукции организма в целом может составить
90-135% (17-стр.48). На основании полученных данных делается вывод о том, что понижение к.п.д. сократительной деятельности мышц является важным механизмом повышения уровня теплопродукции организма. Подчеркивается также тот факт, что речь идет о
понижении к.п.д. мышц не на доли процентов, а в
1,5-3 раза!!! (17-стр.49), аналогичные данные подтверждаются и в других исследованиях (19-стр.13).
Также установлено, что адаптированные к холоду животные по сравнению с адаптированными к теплу имеют более высокий уровень метаболизма, как в тепле, так и в холоде, что выделяет процесс химической терморегуляции в категорию конкурирующего процесса по потреблению кислорода по отношению к мышечной деятельности. Кроме того, проведенные опыты на амбарных крысах определенно указывают на существенное изменение энергетики мышечного сокращения при адаптации животных к холоду, приводящее к увеличению тепловыделения при элементарном сократительном акте (15 – стр.113), что безусловным образом, сказывается на понижении эффективности мышечного сокращения (к.п.д. мышц). При метаболической адаптации к холоду изменяется энергообразование в мышечных волокнах. При этом повышается эффективность отставленного теплообразования за счет снижения к.п.д. химической работы по ресинтезу АТФ (24-стр.128).
При адаптации к холоду происходит интенсификация обмена липидов, возрастает их концентрация в крови и в клетках многих тканей. Липидная перестройка организма вызывается повышением потребности в липидах с одновременным изменением качественного состава липидов. Изменяется соотношение между триглицеридами, фосфолипидами, стеринами, увеличивается доля и концентрация высокомолекулярных (трудноизвлекаемых) жирных кислот, выполняющих роль главного субстрата окисления (24-стр.131), таким образом, повышается плотность жировой ткани и понижается её эластичность и она начинает оказывать ещё большее сопротивление мышечному сокращению. Мышечные волокна не имеют собственных адренорецепторов (преимущественно
a-адренорецепторы), симпатические окончания в мышцах локализованы преимущественно в стенках капилляров и перикапиллярных соединительнотканных образованиях (мышечный жир), расположенных внутри скелетных мышц, а норадреналин индуцирует образование и доставку к мышечным митохондриям свободных жирных кислот.
По современным представлениям, в мышцах млекопитающих выделяют 2 основных типа волокон.
Красные (медленные) – малого диаметра, для обеспечения более эффективного обмена веществ и тепла, они темные, хорошо снабжаются кровью, в них много миоглобина и мало гликогена, к тому же обладают низкой способностью к его расщеплению, содержат большое количество митохондрий, нейтральных жиров и ферментов для обеспечения липидного и окислительного обмена.
Белые (быстрые) – светлые, большого диаметра, содержат мало миоглобина, бедно снабжаются кровью, богаты гликогеном, содержат большое количество гликолитических ферментов и обладают высокой способностью гликогенолиза и гликолиза (превосходящая красные волокна), у них слабая способность к липидному и окислительному обмену. Гликоген мышц служит резервом синтеза АТФ внутри мышечной ткани, для необходимости многократного повышения мышечной активности (
38-стр.158) в результате анаэробного ресинтеза АТФ. При выполнении мышечной работы максимальной напряженности анаэробный путь ресинтеза АТФ обеспечивает на 80-90% общую потребность мышц в АТФ, поскольку гипоксия резко возрастает и сердечно-сосудистая система только ещё начинает увеличивать доставку кислорода к работающим мышцам для удовлетворения их метаболических потребностей (37-стр.10).
Выделяют ещё 2 типа волокон с промежуточными свойствами.
Одни волокна с оксидативно-гликолитическим обменом, а другие обладают обменом, приближающимся к гликолитическому, хотя частично имеют и оксидативный обмен.
Высокоинтенсивные физические нагрузки в пределах 3-х минут повышают активность миозина, Са
2+ и Na
+-K
+-АТФзы, которые осуществляют гидролиз АТФ с высокой скоростью (37-стр.10).
АТФазы
АТФ <--> АДФ+ Ф
н+Н+
где, АДФ – аденозин 5-дифосфат, Ф
н– неорганический фосфат.
Последовательность, в которой происходит анаэробный ресинтез АТФ, при возрастающей потребности в нем, выглядит таким образом (37-стр.10) (Рис.2):
КФК
КФ+АДФ+Н
+ <--> АТФ+Кр
Гликоген+3АДФ+3Ф
н <--> 3АТФ+2лактат-+2Н+
АК
2АДФ <--> АТФ+АМФ
АМФдезаминаза
АМФ+ Н
+ <--> ИМФ+NH4+,
где: КФ – креатинфосфат, К
р - креатин,
КФК – креатинфосфокиназа,
АК – аденилаткиназа,
АМФ – аденозин 5-монофосфат,
ИМФ – инозинмонофосфат.
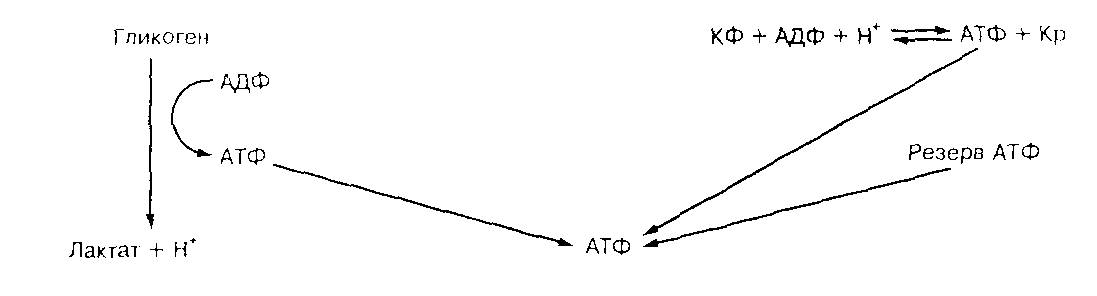
Рис.2 Схема источников анаэробного ресинтеза АТФ.
Распад КФ происходит очень быстро, т.к. он используется как субстрат для ресинтеза АТФ на самой начальной стадии интенсивной физической нагрузки (до 10 с). В то же время с началом интенсивной физической нагрузки активизируется и анаэробные гликолиз и гликогенолиз (37-стр.12-13). Скорость обеспечения ресинтеза АТФ анаэробными путями, является предельной скоростью для осуществления мышечной деятельности высокой мощности и может достигать 2-4 раза большую, чем соответствующую максимальному уровню потребления кислорода (МПК). При выполнении максимальной мышечной деятельности более 10 с, мощность выполняемой работы снижается, а скорость гликолитического и КФ-зависимого ресинтеза АТФ не будет максимальной (37-стр.13). Самые высокие скорости ресинтеза АТФ за счет КФ и гликолитических процессов при физических нагрузках высокой интенсивности проявляются в первые 10 с (Рис.3). При продолжении таких физических нагрузок до 30 с, запасы КФ
истощаются, а скорость гликолитического ресинтеза АТФ будет составлять около 50% скорости ресинтеза в первые 10 с.
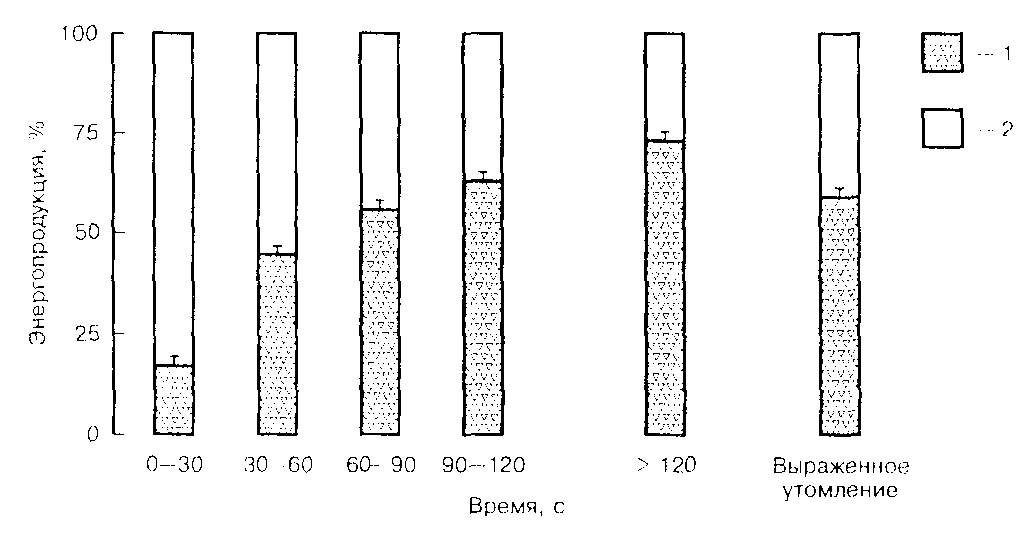
Рис.3 Относительный вклад аэробного (1) и анаэробного (2)
ресинтеза АТФ на протяжении 30-секундных интервалов
предельной (около 3 мин) физической нагрузки
При этом гликолитическое обеспечение АТФ при 30-секундной максимальной физической нагрузке выше в 3…4 раза, чем при обеспечении только за счет КФ-механизма (37-стр.17). Это говорит о том, что тренировочный эффект (за счет развития процессов гликолитического и КФ-механизма ресинтеза АТФ) при тренинге быстроаллюрной лошади будет максимальным, если интервалы максимальных физических нагрузок не будут продолжительностью более 30 с, во избежание развития конкурентоспособности энергообразования аэробного процесса перед анаэробным.
Так в результате тренировки бегунов средний показатель мощности работы при 30-секундном беге с максимальной скоростью увеличился на 60%, а анаэробная продукция АТФ за счет гликолиза выросла на 20%, тогда как вклад энергопродукции за счет КФ и запасов АТФ практически не изменился. Кроме того, имеются литературные данные о том, что под влиянием 4…6 недельной тренировки в велосипедном спринте снизилось содержание красных (медленных) волокон с 57% до 48% и увеличилось содержание белых (быстрых) волокон с 32% до 38% (37-стр.31-32).
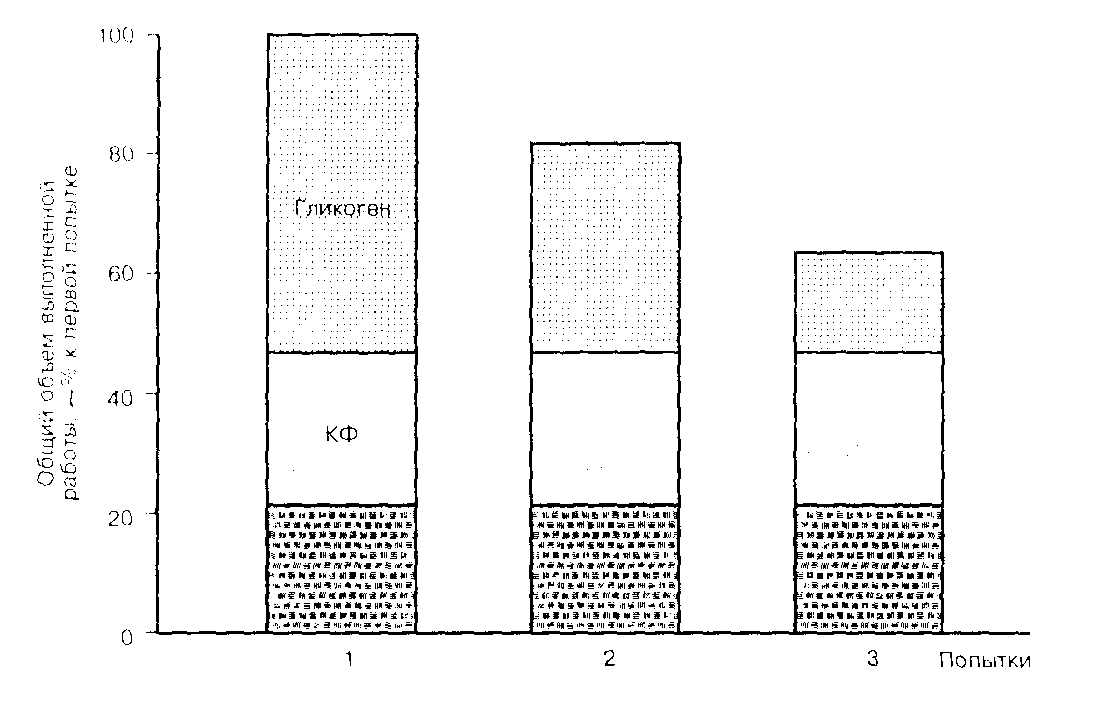
Рис.4 Участие отдельных энергетических механизмов в общей
энергопродукции при трёх максимальных нагрузках продол-
жительностью 30 с каждая с 4-минутными периодами отдыха.
При грамотном применении тренировочных нагрузок, повышение работоспособности скелетных мышц достигается увеличением содержания в них КФ до начала физической нагрузки, а также за счет повышенной способности к его ресинтезу в периоды отдыха. Анаэробные же гликолитические возможности повышаются за счет восстановления истощенных нагрузкой высокой интенсивности запасов в период пониженной мышечной деятельности на фоне высокоуглеводного питания (37-стр.36-37) (Рис.4).
Имеются литературные данные о том, что при адаптации к холоду происходит уменьшение доли белых (быстрых) волокон и увеличение доли красных (медленных) и промежуточных волокон мышечной ткани, а состояние Т-системы (продолжительность следового отрицательного потенциал сопровождающего спайк, пик потенциала действия, ионная проницаемость мембран) белых и красных мышечных волокон стремится к промежуточному состоянию, что выражается проявлением процессов нейрогуморальной координации и дифференциации терморегуляционной активности мышц. Именно для медленных волокон характерны более высокие уровни аэробных процессов, активности окислительных ферментов, концентрации многих энергетических субстратов и миоглобина, лабильность системы окислительного фосфорилирования и её реактивности по отношению к разобщителям (15-стр.85). Так холодовая адаптация белых крыс (4-5 недель)сопровождается заметным повышением активности сукцинатдегидрогеназы (СДГ) в красных волокнах мышцы бедра и особенно в четырехглавой (на 40%). Увеличение активности СДГ происходит почти целиком за счет красных волокон “ядра” мышцы, в которых увеличивается и число митохондрий. “Красное ядро” четырехглавой мышцы контрольных животных составляет 35-38%, а у адаптированных – 47-58% её общего веса, причем разница возрастает с увеличением срока адаптации. Увеличение активности СДГ связывают с повышением мощности сукцинатоксидазной системы, что продиктовано потребностью повышенного окисления сукцината, который в отличие от НАД-зависимых субстратов быстро и многократно увеличивает скорость обмена (20-стр.14-17). Вместе с тем, адаптивные изменения к холоду в мышечных волокнах разных типов остаются малоизученными.
Закономерности различий размеров тела животных одного и того же вида в связи с различиями климатических условий сводятся к тому, что в холодных условиях животные более крупные, отношение единицы поверхности тела к массе тела меньше, размеры выступающих частей тела уменьшены, кроме того, обеспечены более толстыми и эффективными теплоизолирующими средствами (кожный покров, волос, подкожная жировая ткань), чем животные того же вида в теплых климатических зонах (16 – стр.12). Изменения кожного и волосяного покровов организма лошади являются проявлением адаптации вида к условиям внешней среды зоны обитания. Основную функцию в теплозащите от холода путем снижения конвекционных потерь тепла выполняет воздух, находящийся между волосами и в их сердцевинном слое. Стремление к препятствованию потерь тепла заставляет организм лошадей увеличивать число волос, удлинять их длину, развивать сердцевинный слой волос, увеличивать толщину кожного покрова и отложения подкожного жирового слоя (16).
Подключение функциональных резервов для целей адаптации к низким температурам меняет внутреннюю среду организма и включает механизм перестройки для целей выживания (увеличение эритроцитарной массы, объема крови, её компонентов и энергетических депо)(16), понижая полезный потенциал организма на выполнение максимальной физической нагрузки.
Атмосферный воздух содержит промышленные выбросы, но наиболее опасным для организма лошади может стать воздух в конюшне.
Наиболее опасными для лошадей нам представляются соединения азота. Основным источником их появления в воздухе являются разлагающиеся кал, моча и подстилка. В зоотехнической литературе до сих пор присутствует, наряду с другими способами хранения навоза, способ хранения навоза под скотом. Убогость и порочность данной методики кроме как недоумения, ничего более достойного не вызывает.
Все оксиды и закиси азота в соединении с гемоглобином крови составляют метгемоглобин, что приводит к нарушению доставки кислорода к органам и тканям, выделения углекислоты из организма и приводит к приступам асфиксии. Отмечено действие таких соединений на центральную нервную систему. Действие данных соединений безусловным образом тормозит метаболические процессы в организме и вместе с этим, снижает качество полезных признаков животных (резвость, качество и длину прыжка и т.п.).
Аммиак образуется при разложении органических азотсодержащих соединений под действием уреазоактивных анаэробных бактерий. Наиболее высокая концентрация его наблюдается вблизи пола. Аммиак – агрессивный газ. Вызывает сильное раздражение верхних дыхательных путей, конъюнктивы и роговицы глаз, изо рта и носа выделения пенистой жидкости, иногда с примесью крови, нарушение свертываемости крови в результате прямого действия на протромбин, повышение содержания остаточного N
2 в крови за счет быстрого накопления в ней NH3, а также вызывает лейкоцитоз, уменьшение содержания в крови и печени общего и свободного витамина В1. Аммиак рефлекторно вызывает остановки дыхания и кровообращения, могущие привести к смерти. Окись углерода (СО2) усиливает действие аммиака (10). При его вдыхании возможен ожог слизистой оболочки. Уже в малых дозах он парализует деятельность ворсинок дыхательных путей, как щелочь, он подщелачивает кожу и копытный рог, разрыхляя их. При попадании на незащищенный влажный кожный покров, особенно конечности, он под действием кислорода превращается в нитрит – чрезвычайно ядовитое соединение, вызывая цианоз кожных покровов, отставания в росте, отечность и кровоизлияния в миокарде, селезенке, пе


